Structural Biochemistry/Carbohydrates/Carbohydrates and Proteins
Introduction
Many proteins either have carbohydrates added on to them for structural and purposes or use carbohydrates as substrates in cell reactions.
CARBOHYDRATES BINDING MOLECULES AND MALARIA
In a paper titled Carbohydrate Binding Molecules in Malaria Pathology Alan Brown and Matthew K Higgins studied the relationship between glycoproteins and the malaria parasite.
-
 Plasmodium falciparum ring-forms and gametocytes in human blood.
Plasmodium falciparum ring-forms and gametocytes in human blood. -
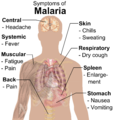 Main symptoms of malaria.
Main symptoms of malaria. -
 The life cycle of malaria parasites in the human body.
The life cycle of malaria parasites in the human body. -
 Anopheles albimanus mosquito feeding on a human arm.
Anopheles albimanus mosquito feeding on a human arm.




Many pathogens, such as bacteria, viruses, and parasites, use carbohydrate-binding molecules to interact with their hosts.Carbohydrates play an important role in the modification and function of proteins.The new insights on pathogenic carbohydrate-binding molecules provide potential therapeutic targets.Glycoproteins play a key role in pathogen binding to their host cells. Malaria-causing parasites, interact with human hepatocytes and erythrocytes. This glycoprotein mediated binding is essential for host cell invasion, movement, avoidance of detection by the immune system, and destruction.
The malaria parasite recognizes multiple sialylated proteins on the erythrocyte surface to bind to the host cell. Two protein families in malaria parasites recognize the sialic acid on the erythrocytes: EBL (erythrocyte-binding like) and RBL (reticulocyte-binding like). Both protein families consists of proteins that interact with different membrane protein and lead to different pathways; their function can be either independent or dependent of sialic acid, which enables the pathogen to overcome the diversities among human population.Both these proteins contain a region called RII. This region is responsible for the ability of the malaria protein to bind to the host cells. EBL and RBL have multiple methods of binding and attack on erythrocytes. This means that it can bind in multiple different ways and avoid destruction by the human immune system.
EBL Family
EBL family is composed of RII region (made of two Duffy-binding like (DBL) domains known as F1 and F2), a C-terminal cysteinerich region and a transmembrane domain.RII region is conservative; it reveals 98% sequence identity across strains from various geographical locations.One of the family members, EBA-175 is better understood by the researchers.
EBA-175
EBA-175 binds to sialylated membrane glycoprotein of erythrocyte, Glycophorin A. Its importance in invasion pathway has been shown through in vitro experiment where antibodies against EBA-175 inhibit binding to Glycophorin A and block invasion.The RII region EBA-175 recognizes and binds to Glycoprotein A, a glycoprotein on the surface of the host cell. Glycoprotein A has specific carbohydrate sequences on its surface that are essential for binding to occur. When the malaria parasite binds to Glycoprotein A, it recognizes sialiac acid on the cell’s surface. This is a good method of attack for the parasite as it is unlikely that sialic acid compositions changes. However, slight differences in sialic acid can prohibit attack by the parasite.
Nevertheless, sialic acid alone is insufficient for pathogenic invasion. For instance, Glycophorin B, which shares 11 conserved sialylated O-linked glycans with Glycophorin A, does not bind EBA-175. In fact, certain orientation of the protein chain is required for optimal binding, where EBA-175 is more likely to directly interact with the protein.
In its crystal form, the two DBL domains of RII region are joined through a three helical linker where two anti-parallel monomers form a dimer. Such structure consists two positively charged channels. Each channels lined with residues from the F1 domain of one monomer and the F2 domain of the other. In solution, though, EBA-175 appears to be concentration dependent in which content ratio of monomer and dimer varies with solute concentration. Thus, it can be proposed that EBA-175 is stable as a monomer and tends to dimerize at high concentrations.
By co-crystallize EBA-175 with Neu5Ac(a2,3)Gal, researchers are able to detect sugar molecule, locating near the channels of dimer, are coordinated by residues of both monomers. This observation aid in the efforts to locate sialic acid binding site and results in the model where EBA-175 assembles around the Glycophorin A ectodomain (glycoproteins on the surface of the membrane and extends into extracellular space) by interactions with both sialic acid and the protein chain.
There are also alternative invasion pathways that involve other members from EBL family. In particular, EBA-140 is devoid of charged patches and residues identified as interacting with sialic acid in EBA-175, while it still binds to Glycophorin C in a sialic acid dependent way. Therefore, each member of EBL family seems to have its own sialic acid binding site distinct from others.
Cytoadhesion and Evasion of Host Detection
Infected erythrocytes become sticky and adhere to uninfected erythrocytes to form clumps known as rosettes. This prevents detection and spleen mediated destruction of infected cells allowing the infection to continue to survive in the body. In Malaria targeted pregnant women, the erythrocytes will accumulate and lead to placental inflammation, possibly causing loss of the child. Antibodies that bind to PfEMP1proteins and prevent adhesion in some forms of Malaria suggests PfEMP1proteins as potential therapeutic targets.PfEMP1. These PfEMP1 proteins are made of DBL (duffy binding-like) domains.
PfEMP1proteins The PfEMP1 proteins contain large-sized ectodomains made from multiple DBL and CIDR domains. Different PfEMP1 proteins will interact with different receptors. Among PfEMP1 proteins, VAR2CSA is the best understood by the researchers.PfEMP1s switch rapidly to avoid detection in the body. These interact with various proteins (CD36 and ICAM-1) as well as carbohydrates. One of these PfEMPS1 is VAR2CSA.
VAR2CSA
The infected erythrocytes by VAR2SCA become specifically avid for 4-sulphated chondroitin sulphate A (CSA) molecules. CSA molecules, especially those that are closely related glycosaminoglycan carbohydrates, show little ability to prevent interactions with VAR2CSA. In addition, the protein chain of CSA molecule does not contribute to the specificity of recognition of the molecule.Structurally, Four out of the six DBL domains from VAR2CSA bind to CSA in isolation. The DBL3X structure has an a-helical structure similar to the EBA-175 DBL domains, only with longer loops.
When sulphate ions or disaccharides are present, a loop on the concave surface of the domain will form a positively charged patch and sulphate-binding pocket crucial for CSA binding. Different form EBA-175, VAR2CSA has longer, often disordered, loops and is more versatile. The versatility of VAR2CSA hints that VAR2CSA has a longer length of exposure to immune system in comparison to EBA-175 during invasion process. The structure of the DBL6e domain is similar toDBL3X, but it does not have the sulphate-binding pocket or positively charged patch. It binds to CSA molecule through positive charges on a distinct surface of the domain, a mechanism that shows more versatility than DBL3X. Therefore, it is more likely to be a more exposed part of the intact ectodomain during cell invasion.
The two domains have distinct surface features to interact with CAS molecules. Moreover, they do not have the CSA-binding specificity; rather their affinity for carbohydrates increases with the charge of the carbohydrate. In addition, the ectodomain shows strong ligand specificity where the ectodomain acts as a compacted monomer distinct fromEBA-175.
Conclusion
Carbohydrate binding molecules are crucial in the life cycle of Malaria pathogen. Both EBA-175 and VAR2CSA use the same DBL domain in binding carbohydrates where the specificity of binding requires multiple domains. EBA-175 needs both protein and carbohydrate components of a dimeric receptor for specificity whereas VAR2CSA requires only carbohydrate for specificity; EBA-175 uses “carbohydrate binding surfaces lining the grooves at dimerization interfaces” whereas VAR2CSA uses “multiple domains from a single monomer to generate a specific binding pocket”. Studies of the carbohydrate-binding proteins and deeper understanding of their structures can provide useful information on pathogenic pathway as well as potential therapeutic targets in the future.