Earlier the name "carbohydrate" was used in chemistry for any compound with the formula Cm(H2O)n. Following this definition, some chemists considered formaldehyde CH2O to be the simplest carbohydrate,
while others claimed that title for glycolaldehyde. Today the term is generally understood in the biochemistry sense, which excludes compounds with only one or two carbons atoms.
Natural saccharides are generally built of simple carbohydrates called monosaccharides with general formula (CH2O)n where n is three or more. A typical monosaccharide has the structure H-(CHOH)x(C=O)-(CHOH)y-H, that is, an aldehyde or ketone with many hydroxyl groups added, usually one on each carbon atom that is not part of the aldehyde or ketone functional group. Examples of monosaccharides are glucose, fructose, and glyceraldehyde. However, some biological substances commonly called "monosaccharides" do not conform to this formula (e.g., uronic acids and deoxy-sugars such as fucose), and there are many chemicals that do conform to this formula but are not considered to be monosaccharides (e.g., formaldehyde CH2O and inositol (CH2O)6).
The open-chain form of a monosaccharide often coexists with a heterocyclic compound|closed ring form where the aldehyde/ketone carbonyl group carbon (C=O) and hydroxyl group (-OH) react forming a hemiacetal with a new C-O-C bridge.
Monosaccharides can be linked together into what are called polysaccharides (or oligosaccharides) in a large variety of ways. Many carbohydrates contain one or more modified monosaccharide units that have had one or more groups replaced or removed. For example, deoxyribose, a component of DNA, is a modified version of ribose; chitin is composed of repeating units of N-acetylglucosamine, a nitrogen-containing form of glucose.[1]
Monosaccharide Conformation:
Pyranose and furanose forms can exist in different conformers and one can interconvert between the different conformations if an energy penalty is met. For the furanose system there are two possible conformers: Twist (T) and Envelope (E). In the pyranose system four conformers are possible: Chair (C), Boat (B), Skew (S), Half-Chair (H) or Envelope (E). In all cases there are four or more atoms that make up a plane. In order to define which atoms are above and below the plane one must orient the molecule so that the atoms are numbered clockwise when looking form the top. Atoms above the plane are prefixed as a superscript and atoms below the plane are suffixed as a subscript. If the ring oxygen is above or below the plane it must be prefixed or suffixed appropriately
Conformational Analysis
Chair conformation of six membered rings have a dihedral angle of 60° between adjacent substituents thus making it the most stable conformer. Since there are two possible chair conformation steric and stereoelectronic effects such as the anomeric effect, 1,3 diaxial interactions, dipoles and intramolecular hydrogen bonding must be taken into consideration when looking at relative energies. Conformations with 1,3 diaxial interactions are usually disfavored due to steric congestion and can shift equilibrium to the other chair form (example: 1C4 to 4C1). The size of the substituents greatly affects this equilibrium. However, intramolecular hydrogen bonding can be an example of a stabilizing 1,3 diaxial interaction. Dipoles also play a role in conformer stability, aligned dipoles lead to an increase in energy while opposed dipoles lead to a lowering of energy hence a stabilizing effect, this can be complicated by solvent effects. Polar solvents tend to stabilize aligned dipoles. All interaction must be taken into account when determining a preferred conformation
Conformations of five membered rings are limited to two, envelope and twist. The envelope conformation has four atoms in a plane while the twist form only has three. In the envelope form two different scenarios can be envisioned; one where the ring oxygen is in the four atom plane and one where it is puckered above or below the plane. When the ring oxygen is not in the plane the substituents eclipse and when it is in the plane torsional strain is relieved. Conformational analysis for the twist form is similar thus leading to the two forms being very close in energy.[2]
Anomers are diastereoisomers of glycosides, hemiacetals or related cyclic forms of sugars, or related molecules differing in configuration only at C-1. When the stereochemistry of the first carbon matches the stereochemistry of the last stereogenic center the sugar is the α-anomer when they are opposite the sugar is the β-anomer.In organic chemistry, the anomeric effect or Edward-Lemieux effect is a stereoelectronic effect that describes the tendency of heteroatomic substituents adjacent to a heteroatom within a cyclohexane ring to prefer the axial orientation instead of the less hindered equatorial orientation that would be expected from steric considerations. This effect was originally observed in pyranose rings by J. T. Edward in 1955; at that time, N.-J. Chii and R. U. Lemieux began to study the anomerization equilibria of the fully acetylated derivatives of several aldohexopyranoses. The term "anomeric effect" was introduced in 1958.The anomeric effect got its name from the term used to designate the C-1 carbon of a pyranose, the anomeric carbon. Isomers that differ only in the configuration at the anomeric carbon are called anomers.[3]
The α- and β-anomers of D-glucopyranose.
The anomers of glucopyranose are diastereomers, with the beta anomer on the right having an OH group pointing up equatorially in the lower right-hand corner of the figure, and the alpha anomer on the left having that OH group pointing down axially.
The anomeric effect can be generalized to any system with the general formula R–Y–C–Z, where Y is an atom with one or more electronic lone pairs, and Z is an electronegative atom. The magnitude of the anomeric effect is estimated at about 1–2 kcal/mol in the case of sugars. In this general case, the molecule need not be cyclic. For example, a small molecule that exhibits the anomeric effect and that is often used for theoretical studies is dimethoxymethane. In the case of dimethoxyethane the gauche, gauche conformation is about 3–5 kcal/mol lower in energy (more stable) than the trans, trans conformation—this is about two times as big as the effect in sugars because there are two rotatable bonds that are affected.[4]
Anomeric Effect
Anomers can be interconverted through a process know as mutarotation. The anomeric effect more accurately called the endo-anomeric effect is the propensity for heteroatoms at C-1 to be oriented axially. This is counter intuitive as one would expect the equatorially anomer to be the thermodynamic product. This effect has been rationalized through dipole-dipole repulsion and n-σ* arguments.[5]
Reverse Anomeric Effect
The reverse anomeric effect, proposed in 1965 by R. U. Lemieux, is the tendency for electropositive groups at the anomeric position to be oriented equatorially. Original publication reported this phenomenon with N-(2,3,4,6-tetra-O-acetyl-α-D-glucopyranosyl)-4-methylpyridinium bromide. However, further studies have shown the effect to be a solvation and steric issue. It is accepted that there is no generalized reverse anomeric effect.[6]
Hydroxymethyl Conformation
Rotation around the C-5/C-6 bond is described by the angle ω. Three possible staggered conformations possible; gauche-trans (gt), gauche-gauche (gg), trans-gauche(tg). The name indicates the interaction between O-5 and OH-6 first followed by the interaction between OH-6 and C-4.[7]
In addition to the factors affecting monosaccaride residues, conformational analysis of oligosaccharides and polysaccharides requires consideration additional factors.
Exo-Anomeric Effect
Exo-anomeric effect is similar to the endo-anomeric effect. The difference being that the lone pair being donated is coming from the substituent at C-1. However, since the substituent can be either axial or equatorial there are two types of exo-anomeric effects, one from axial glycosides and one from equatorial glycosides as long as the donating orbital is anti-periplanar to the accepting orbital.
Glycosidic Torsion Angles
Three angles are described by φ, ψ and ω (in the case of glycosidic linkages via O-6). Steric considerations and anomeric effects need to be taken into consideration when looking at preferred angles.
α-D-Glucopyranose
Pyranose is a collective term for carbohydrates that have a chemical structure that includes a six-membered ring consisting of five carbon atoms and one oxygen atom. The name derives from its similarity to the oxygen heterocycle pyran, but the pyranose ring does not have double bonds. A pyranose in which the anomeric OH at C(1) has been converted into an OR group is called a pyranoside.[8]
Formation of Pyranose
The pyranose ring is formed by the reaction of the hydroxyl group on carbon 5 (C-5) of a sugar with the aldehyde at carbon 1. This forms an intramolecular hemiacetal. If reaction is between the C-4 hydroxyl and the aldehyde, a furanose is formed instead.
Formation of pyranose hemiacetal and representations of beta-D-glucopyranoseHaworth Projection of beta-D-glucopyranose
Hermann Emil Fischer won the Nobel Prize in Chemistry (1902) for his work in determining the structure of the D-aldohexoses. However, the linear, free-aldehyde structures that Fischer proposed represent a very minor percentage of the forms that hexose sugars adopt in solution. It was Edmund Hirst and Clifford Purves, in the research group of Walter Haworth, who conclusively determined that the hexose sugars preferentially form a pyranose, or six-membered, ring. Haworth drew the ring as a flat hexagon with groups above and below the plane of the ring – the Haworth projection.
A further refinement to the conformation of pyranose rings came when Sponsler and Dore (1926) realized that Sachse’s mathematical treatment of six-membered rings could be applied to their X-ray structure of cellulose.It was determined that the pyranose ring is puckered, to allow all of the carbon atoms of the ring to have close to the ideal tetrahedral geometry.
This puckering leads to a total of 38 distinct basic pyranose conformations: 2 chairs, 6 boats, 6 skew-boats, 12 half-chairs, and 12 envelopes.[10]
Relative energy of beta-D-glucopyranose conformers
These conformers can interconvert with one another; however, each form may have very different relative energy, so a significant barrier to interconversion may be present. The energy of these conformations can be calculated from quantum mechanics; an example of possible glucopyranose interconversions is given.[11]
The conformations of the pyranose ring are superficially similar to that of the cyclohexane ring. However, the specific nomenclature of pyranoses includes reference to the ring oxygen, and the presence of hydroxyls on the ring have distinct effects on its conformational preference. There are also conformational and stereochemical effects specific to the pyranose ring.
To name conformations of pyranose, first the conformer is determined. The common conformers are similar to those found in cyclohexane, and these form the basis of the name. Common conformations are Chair (C), Boat (B), Skew (S), Half-Chair (H) or Envelope (E). The ring atoms are then numbered; the anomeric, or hemiacetal, carbon is always 1. Oxygen atoms in the structure are, in general, referred to by the carbon atom they are attached to in the acyclic form, and designated O. Then:
Position the ring so that, if looking at the top face, the atoms are numbered clockwise. 4 (or 5, in the case of an envelope) atoms will be in a plane
Atoms above the plane are written before the conformer label, as a superscript
Atoms below the plane are written following the conformer label, as a subscript
"Family tree" of the D -aldoses. By appending CH-OH groups to extend the basic framework, so more sugar can be derived (from trio with three C-hexoses to six carbon atoms). Here, the rotation of polarized light with (+) and) (specified -. ( 1 ) D -(+)- glyceraldehyde , ( 2 ) D -(-)- erythrose , ( 2b ) D -(-)- threose , ( 3a ) D -(-)- ribose , ( 3b ) D - ( -) - arabinose , ( 3c ) D -(+)- xylose , ( 3d ) D -(-)- lyxose , ( 4a ) D -(+)- allose , ( 4b ) D -(+)- altrose ; ( 4c ) D -(+)- glucose , ( 4d ) D -(+)- mannose , ( 4e ) D -(-)- gulose , ( 4f ) D -(-)- idose ; ( 4g ) D - (+ ) - galactose , ( 4 h ) D -(+)- talose
Monosaccharides are the simplest carbohydrates in that they cannot be hydrolyzed to smaller carbohydrates. They are aldehydes or ketones with two or more hydroxyl groups. The general chemical formula of an unmodified monosaccharide is (C•H2O)n, literally a "carbon hydrate." Monosaccharides are important fuel molecules as well as building blocks for nucleic acids. The smallest monosaccharides, for which n = 3, are dihydroxyacetone and D- and L-glyceraldehyde.
"Family tree" of the D -ketose. By appending CH-OH groups to extend the basic framework, so more sugar can be derived (from Ketotriosen with three C-ketohexoses to six carbon atoms). ( a ) dihydroxyacetone , ( 2 ) D - Erythrulose , ( 3 ) D - ribulose , ( 3b ) D - xylulose , ( 4 ) D - psicose , ( 4b ) D - fructose , ( 4c ) D - sorbose , ( 4d ) D - tagatose
A triose is a monosaccharide containing three carbon atoms. There are only two trioses, an aldotriose (glyceraldehyde) and a ketotriose (dihydroxyacetone). Trioses are important in respiration. Namely, lactic acid and pyruvic acid are derived from aldotriose and ketotriose, respectively.[14]
The D-aldotriose is D-Glyceraldehyde
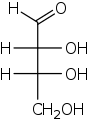
A tetrose is a monosaccharide with 4 carbon atoms. They have either an aldehyde functional group in position 1 (aldotetroses) or a ketone functional group in position 2 (ketotetroses).The aldotetroses have two chiral centers ("asymmetric carbon atoms") and so 4 different stereoisomers are possible. There are two naturally occurring stereoisomers, the enantiomers of erythrose and threose having the D configuration but not the L enantiomers. The ketotetroses have one chiral center and, therefore, two possible stereoisomers: erythrulose (L- and D-form). Again, only the D enantiomer is naturally occurring.[15]
D-Erythrose
D-Threose
D-Erythrulose
Pentose
A pentose is a monosaccharide with five carbon atoms. Pentoses are organized into two groups. Aldopentoses have an aldehyde functional group at position 1. Ketopentoses have a ketone functional group in position 2 or 3. The aldehyde and ketone functional groups in these carbohydrates react with neighbouring hydroxyl functional groups to form intramolecular hemiacetals and hemiketals, respectively. The resulting ring structure is related to furan, and is termed a furanose. The ring spontaneously opens and closes, allowing rotation to occur about the bond between the carbonyl group and the neighbouring carbon atom — yielding two distinct configurations (α and β). This process is termed mutarotation.
Ribose is a constituent of RNA, and the related deoxyribose of DNA.
A polymer composed of pentose sugars is called a pentosan.[16]
Aldopentoses
The aldopentoses have three chiral centers and therefore eight different stereoisomers are possible.
Hexose
In Biochemistry, a hexose is a monosaccharide with six carbon atoms, having the chemical formula C6H12O6. Hexoses are classified by functional group, with aldohexoses having an aldehyde at position 1, and ketohexoses having a ketone at position 2. [17]
Aldohexoses
The aldohexoses have four chiral centres for a total of 16 possible aldohexose stereoisomers (24). The D/L configuration is based on the orientation of the hydroxyl at position 5, and does not refer to the direction of optical activity.
The eight D-aldohexoses are[18]:
Of these D-isomers, all except D-altrose are naturally occurring. L-Altrose, however, has been isolated from strains of the bacterium Butyrivibrio fibrisolvens.
A classical jingle to remember the hexoses is "All atruists gladly make gum in gallon tank".
Cyclic hemiacetals
It has been known since 1926 that 6-carbon aldose sugars form cyclic hemiacetals.[19] The diagram below shows the hemiacetal forms for D-glucose and D-mannose.
The numbered carbons in the open-chain forms correspond to the same numbered carbons in the hemiacetal forms. The formation of the hemiacetal causes carbon number 1, which is symmetric in the open-chain form, to become asymmetric in the cyclic version. This means that both glucose and mannose (as well as all the other aldohexoses) each have two cyclic forms. In solution, both of these exist in equilibrium with the open-chain form. The open-chain form, however, does not crystallize. Hence the two cyclic forms become separable when they are crystallized. For example, D-glucose forms an alpha crystal that has specific rotation of +112° and melting point of 146 °C, as well as a beta crystal that has specific rotation of +19° and melting point of 150 °C.[19][20]
Ketohexoses
The ketohexoses have 3 chiral centres and therefore eight possible stereoisomers (23). Of these, only the four D-isomers are known to occur naturally: [21]
Only the naturally occurring hexoses are capable of being fermented by yeasts.
Mutarotation:
The aldehyde and ketone functional groups in these carbohydrates react with neighbouring hydroxyl functional groups to form intramolecularhemiacetals and hemiketals, respectively. The resulting ring structure is related to pyran, and is termed a pyranose. The ring spontaneously opens and closes, allowing rotation to occur about the bond between the carbonyl group and the neighbouring carbon atom, yielding two distinct configurations (α and β). This process is termed mutarotation. Hexose sugars can form dihexose sugars with a condensation reaction to form a 1,6-glycosidic bond.[22]
Heptose
A heptose is a monosaccharide with seven carbon atoms.
They have either an aldehyde functional group in position 1 (aldoheptoses) or a ketone functional group in position 2 (ketoheptoses).
There are few examples of C-7 sugars in nature, among which are[23]:
Sedoheptulose or D-altro-heptulose (a ketose)
Mannoheptulose, found in avocados
L-glycero-D-manno-heptose (an aldose).
Ketoheptoses have 4 chiral centers, whereas aldoheptoses have 5[24].
The three-dimensional structure of a monosaccharides in cyclic form is usually represented by its Haworth projection. In this diagram, the α-isomer has the OH- of the anomeric carbon below the plane of the carbon atoms, and the β-isomer has the OH- of the anomeric carbon above the plane. Pyranoses typically adopt a chair conformation, similar to cyclohexane. In this conformation the α-isomer has the OH- of the anomeric carbon in an axial position, whereas the β-isomer has the OH- of the anomeric carbon in equatorial position.[25]
α-D-Glucopyranose
β-D-Glucopyranose
What is Mutarotation? Mutarotation is the change in the optical rotation that occurs by epimerization (that is the change in the equilibrium between two epimers, when the corresponding stereocenters interconvert). Cyclic sugars show mutarotation as α and β anomeric forms interconvert.[26]
The optical rotation of the solution depends on the optical rotation of each anomer and their ratio in the solution.
Mutarotation was discovered by Augustin-Pierre Dubrunfaut in 1846, when he noticed that the specific rotation of aqueous sugar solution changes with time. [27][28]
Measurement
Interconversion of α and β anomers
The α and β anomers are diastereomers of each other and usually have different specific rotations. A solution or liquid sample of a pure α anomer will rotate plane polarised light by a different amount and/or in the opposite direction than the pure β anomer of that compound. The optical rotation of the solution depends on the optical rotation of each anomer and their ratio in the solution.
For example if a solution of β-D-glucopyranose is dissolved in water, its specific optical rotation will be +18.7. Over time, some of the β-D-glucopyranose will undergo mutarotation to become α-D-glucopyranose, which has an optical rotation of +112.2. Thus the rotation of the solution will increase from +18.7 to an equilibrium value of +52.5 as some of the β form is converted to the α form. The equilibrium mixture is actually about 64% of β-D-glucopyranose and about 36% of α-D-glucopyranose, though there are also with traces of the other forms including furanoses and open chained form.[29]
The observed optical rotation of the sample is the weighted sum of the optical rotation of each anomer weighted by the amount of that anomer present. Therefore one can use a polarimeter to measure the rotation of a sample and then calculate the ratio of the two anomers present from the enantiomeric excess, as long as one knows the rotation of each pure anomer. One can monitor the mutarotation process over time or determine the equilibrium mixture by observing the optical rotation and how it changes.
A monosaccharide often switches from the acyclic (open-chain) form to a cyclic form, through a nucleophilic addition reaction between the carbonyl group and one of the hydroxyls of the same molecule. The reaction creates a ring of carbon atoms closed by one bridging oxygen atom. The resulting molecule has an hemiacetal or hemiketal group, depending on whether the linear form was an aldose or a ketose. The reaction is easily reversed, yielding the original open-chain form.
In these cyclic forms, the ring usually has 5 or 6 atoms. These forms are called furanoses and pyranoses, respectively — by analogy with furan and pyran, the simplest compounds with the same carbon-oxygen ring (although they lack the double bonds of these two molecules). For example, the aldohexose glucose may form a hemiacetal linkage between the hydroxyl on carbon 1 and the oxygen on carbon 4, yielding a molecule with a 5-membered ring, called glucofuranose. The same reaction can take place between carbons 1 and 5 to form a molecule with a 6-membered ring, called glucopyranose. Cyclic forms with a 7-atom ring (the same of oxepane), rarely encountered, are called septanoses.For many monosaccharides (including glucose), the cyclic forms predominate, in the solid state and in solutions, and therefore the same name commonly is used for the open- and closed-chain isomers. Thus, for example, the term "glucose" may signify glucofuranose, glucopyranose, the open-chain form, or a mixture of the three.
Cyclization creates a new stereogenic center at the carbonyl-bearing carbon The -OH group that replaces the carbonyl's oxygen may end up in two distinct positions relative to the ring's midplane. Thus each open-chain monosaccharide yields two cyclic isomers (anomers), denoted by the prefixes 'α-' and 'β-'. The molecule can change between these two forms by a process called mutarotation, that consists in a reversal of the ring-forming reaction followed by another ring formation.[30]
Conversion between the furanose, acyclic, and pyranose forms of D-glucose.Pyranose forms of some pentose sugars.Pyranose forms of some hexose sugars.
There are two different types of disaccharides: reducing disaccharides, in which one monosaccharide, the reducing sugar, still has a free hemiacetal unit; and non-reducing disaccharides, in which the components bond through an acetal linkage between their anomeric centers and neither monosaccharide has a free hemiacetal unit. Cellobiose and maltose are examples of reducing disaccharides. Sucrose and trehalose are examples of non-reducing disaccharides.
Formation[31]
Disaccharides are formed when two monosaccharides are joined together and a molecule of water is removed. For example; milk sugar (lactose) is made from glucose and galactose whereas the sugar from sugar cane and sugar beets (sucrose) is made from glucose and fructose.
The two monosaccharides are bonded via a dehydration reaction (also called a condensation reaction or dehydration synthesis) that leads to the loss of a molecule of water and formation of a glycosidic bond.[32]
Properties
The glycosidic bond can be formed between any hydroxyl group on the component monosaccharide. So, even if both component sugars are the same (e.g., glucose), different bond combinations (regiochemistry) and stereochemistry (alpha- or beta-) result in disaccharides that are diastereoisomers with different chemical and physical properties.
Depending on the monosaccharide constituents, disaccharides are sometimes crystalline, sometimes water-soluble, and sometimes sweet-tasting and sticky-feeling.[33]
Sucrose is the organic compound commonly known as table sugar and sometimes called saccharose. A white, odorless, crystalline powder with a sweet taste, it is best known for its role in human nutrition. The molecule is a disaccharide derived from glucose and fructose with the molecular formula C12H22O11. About 150,000,000 tonnes (metric tons) are produced annually.
Lactulose is a synthetic sugar used in the treatment of constipation[1] and hepatic encephalopathy, a complication of liver disease. It is a disaccharide (double-sugar) formed from one molecule each of the simple sugars (monosaccharides) fructose and galactose. The commercial syrup used for treatment of constipation is dyed yellow-orange. It is produced commercially by isomerization of lactose.
Lactose was discovered in milk in 1619 by Fabriccio Bartoletti, and identified as a sugar in 1780 by Carl Wilhelm Scheele. Lactose is a disaccharide sugar that is found most notably in milk and is formed from galactose and glucose. Lactose makes up around 2~8% of milk (by weight), although the amount varies among species and individuals. It is extracted from sweet or sour whey. The name comes from lac, the Latin word for milk, plus the -ose ending used to name sugars. It has a formula of C12H22O11.
Maltose (English pronunciation: /ˈmɒltoʊz/), or malt sugar, is a disaccharide formed from two units of glucose joined with an α(1→4)bond. The isomer "isomaltose" has two glucose molecules linked through an α(1→6) bond. Maltose is the second member of an important biochemical series of glucose chains. Maltose is the disaccharide produced when amylase breaks down starch. It is found in germinating seeds such as barley as they break down their starch stores to use for food.
The addition of another glucose unit yields maltotriose; further additions will produce dextrins (also called maltodextrins) and eventually starch (glucose polymer).
Maltose can be broken down into two glucose molecules by hydrolysis. In living organisms, the enzyme maltase can achieve this very rapidly. In the laboratory, heating with a strong acid for several minutes will produce the same result. Isomaltose is broken by isomaltase.
The production of maltose from germinating cereals, such as barley, is an important part of the brewing process. When barley is malted, it is brought into a condition in which the concentration of maltose-producing amylases has been maximized. Mashing is the process by which these amylases convert the cereal's starches into maltose. Metabolism of maltose by yeast during fermentation then leads to the production of ethanol and carbon dioxide.[35]
Trehalose, also known as mycose or tremalose, is a natural alpha-linked disaccharide formed by an α,α-1,1-glucoside bond between two α-glucose units. In 1832, H.A.L. Wiggers discovered trehalose in an ergot of rye, and in 1859 Marcellin Berthelot isolated it from trehala manna, a substance made by weevils, and named it trehalose. It can be synthesised by fungi, plants, and invertebrate animals. It is implicated in anhydrobiosis — the ability of plants and animals to withstand prolonged periods of desiccation. It has high water retention capabilities, and is used in food and cosmetics. The sugar is thought to form a gel phase as cells dehydrate, which prevents disruption of internal cell organelles, by effectively splinting them in position. Rehydration then allows normal cellular activity to be resumed without the major, lethal damage that would normally follow a dehydration/rehydration cycle. Trehalose has the added advantage of being an antioxidant. Extracting trehalose used to be a difficult and costly process, but, recently, the Hayashibara company (Okayama, Japan) confirmed an inexpensive extraction technology from starch for mass production. Trehalose is currently being used for a broad spectrum of applications.[36]
Cellobiose is a disaccharide with the formula [HOCH2CHO(CHOH)3]2O. The molecule is derived from the condensation of two glucose molecules linked in a β(1→4) bond. It can be hydrolyzed by bacteria or cationic ion exchange resins to give glucose. Cellobiose has eight free alcohol (COH) groups and three ether linkages, which give rise to strong inter- and intra-molecular hydrogen bonds
It can be obtained by enzymatic or acidic hydrolysis of cellulose and cellulose rich materials such as cotton, jute, or paper. Cellulose is a polymer of glucose units linked by β(1→4) bonds.
Treatment of cellulose with acetic anhydride and sulfuric acid, gives cellobiose octoacetate, which cannot engage in hydrogen bonding and is soluble in nonpolar organic solvents.[37]
Maltose and cellobiose are hydrolysis products of the polysaccharides, starch and cellulose, respectively.
Polysaccharides are polymeric carbohydrate structures, formed of repeating units (either mono- or di-saccharides) joined together by glycosidic bonds. These structures are often linear, but may contain various degrees of branching. Polysaccharides are often quite heterogeneous, containing slight modifications of the repeating unit. Depending on the structure, these macromolecules can have distinct properties from their monosaccharide building blocks. They may be amorphous or even insoluble in water.
Polysaccharides have a general formula of Cx(H2O)y where x is usually a large number between 200 and 2500. Considering that the repeating units in the polymer backbone are often six-carbon monosaccharides, the general formula can also be represented as (C6H10O5)n where 40≤n≤3000.
Starch or amylum is a carbohydrate consisting of a large number of glucose units joined together by glycosidic bonds. This polysaccharide is produced by all green plants as an energy store. It is the most important carbohydrate in the human diet and is contained in such staple foods as potatoes, wheat, maize (corn), rice, and cassava.
Pure starch is a white, tasteless and odorless powder that is insoluble in cold water or alcohol. It consists of two types of molecules: the linear and helical amylose and the branched amylopectin. Depending on the plant, starch generally contains 20 to 25% amylose and 75 to 80% amylopectin. Glycogen, the glucose store of animals, is a more branched version of amylopectin.
Starch is processed to produce many of the sugars in processed foods. When dissolved in warm water, it can be used as a thickening, stiffening or gluing agent, giving wheatpaste.
Amylose is made up of α(1→4) bound glucose molecules. The carbon atoms on glucose are numbered, starting at the aldehyde (C=O) carbon, so, in amylose, the 1-carbon on one glucose molecule is linked to the 4-carbon on the next glucose molecule (α(1→4) bonds). The structural formula of amylose is pictured at right. The number of repeated glucose subunits (n) is usually in the range of 300 to 3000, but can be many thousands.
There are three main forms of amylose chains can take. It can exist in a disordered amorphous conformation or two different helical forms. It can bind with itself in a double helix (A or B form), or it can bind with another hydrophobic guest molecule such as iodine, a fatty acid, or an aromatic compound. This is known as the V form and is how amylopectin binds to amylose to form starch. Within this group, there are many different variations. Each is notated with V and then a subscript indicating the number of glucose units per turn. The most common is the V6 form, which has six glucose units a turn. V8 and possibly V7 forms exist as well. These provide an even larger space for the guest molecule to bind.
This linear structure can have some rotation around the phi and psi angles, but, for the most part, bound glucose ring oxygens lie on one side of the structure. The α(1→4) structure promotes the formation of a helix structure, making it possible for hydrogen bonds form between the oxygen atoms bound at 2-carbon of one glucose molecule and the 3-carbon of the next glucose molecule.
Amylopectin is a soluble polysaccharide and highly branched polymer of glucose found in plants. It is one of the two components of starch, the other being amylose.
Glucose units are linked in a linear way with α(1→4) glycosidic bonds. Branching takes place with α(1→6) bonds occurring every 24 to 30 glucose units. In contrast, amylose contains very few α(1→6) bonds, which causes it to be hydrolyzed more slowly but have higher density.
Its counterpart in animals is glycogen, which has the same composition and structure, but with more extensive branching that occurs every 8 to 12 glucose units.
Plants store starch within specialized organelles called amyloplasts. When energy is needed for cell work, the plant hydrolyzes the starch, releasing the glucose subunits. Humans and other animals that eat plant foods also use amylase, an enzyme that assists in breaking down amylopectin.
Starch is made of about 70% amylopectin by weight, though it varies depending on the source (higher in medium-grain rice to 100% in waxy rice, waxy potato starch, and waxy corn, and lower in long-grain rice, amylomaize, and russet potato, for example). Amylopectin is highly branched, being formed of 2,000 to 200,000 glucose units. Its inner-chains are formed of 20-24 glucose subunits.[41]
Glycogen is the molecule that functions as the secondary long-term energy storage in animal and fungal cells. It is made primarily by the liver and the muscles, but can also be made by glycogenesis within the brain and stomach.
Glycogen is the analogue of starch, a less branched glucose polymer in plants, and is commonly referred to as animal starch, having a similar structure to amylopectin. Glycogen is found in the form of granules in the cytosol in many cell types, and plays an important role in the glucose cycle. Glycogen forms an energy reserve that can be quickly mobilized to meet a sudden need for glucose, but one that is less compact than the energy reserves of triglycerides (lipids). In the liver hepatocytes, glycogen can compose up to 8% of the fresh weight (100–120 g in an adult) soon after a meal. Only the glycogen stored in the liver can be made accessible to other organs. In the muscles, glycogen is found in a low concentration (1% to 2% of the muscle mass). However, the amount of glycogen stored in the body - especially within the red blood cells,liver, and muscles - mostly depends on physical training, basal metabolic rate, and eating habits such as intermittent fasting. Small amounts of glycogen are found in the kidneys, and even smaller amounts in certain glial cells in the brain and white blood cells. The uterus also stores glycogen during pregnancy to nourish the embryo.
File:AmylopectinAmylopectinAmylopectin. The main chain is the same as in amylose
A strand of cellulose (conformation Iα), showing the hydrogen bonds (dashed) within and between cellulose molecules.
Cellulose
The structural component of plants are formed primarily from cellulose. Wood is largely cellulose and lignin, while paper and cotton are nearly pure cellulose. Cellulose is a polymer made with repeated glucose units bonded together by beta-linkages. Humans and many other animals lack an enzyme to break the beta-linkages, so they do not digest cellulose. Certain animals can digest cellulose, because bacteria possessing the enzyme are present in their gut. The classic example is the termite.Cellulose has no taste, is odourless, is hydrophilic with the contact angle of 20–30, is insoluble in water and most organic solvents, is chiral and is biodegradable. It can be broken down chemically into its glucose units by treating it with concentrated acids at high temperature.
Cellulose is derived from D-glucose units, which condense through β(1→4)-glycosidic bonds. This linkage motif contrasts with that for α(1→4)-glycosidic bonds present in starch, glycogen, and other carbohydrates. Cellulose is a straight chain polymer: unlike starch, no coiling or branching occurs, and the molecule adopts an extended and rather stiff rod-like conformation, aided by the equatorial conformation of the glucose residues. The multiple hydroxyl groups on the glucose from one chain form hydrogen bonds with oxygen molecules on the same or on a neighbor chain, holding the chains firmly together side-by-side and forming microfibrils with high tensile strength. This strength is important in cell walls, where the microfibrils are meshed into a carbohydrate matrix, conferring rigidity to plant cells.
Compared to starch, cellulose is also much more crystalline. Whereas starch undergoes a crystalline to amorphous transition when heated beyond 60-70 °C in water (as in cooking), cellulose requires a temperature of 320 °C and pressure of 25 MPa to become amorphous in water.
Several different crystalline structures of cellulose are known, corresponding to the location of hydrogen bonds between and within strands. Natural cellulose is cellulose I, with structures Iα and Iβ. Cellulose produced by bacteria and algae is enriched in Iα while cellulose of higher plants consists mainly of Iβ. Cellulose in regenerated cellulose fibers is cellulose II. The conversion of cellulose I to cellulose II is not reversible, suggesting that cellulose I is metastable and cellulose II is stable. With various chemical treatments it is possible to produce the structures cellulose III and cellulose IV.
Many properties of cellulose depend on its chain length or degree of polymerization, the number of glucose units that make up one polymer molecule. Cellulose from wood pulp has typical chain lengths between 300 and 1700 units; cotton and other plant fibers as well as bacterial celluloses have chain lengths ranging from 800 to 10,000 units.[10] Molecules with very small chain length resulting from the breakdown of cellulose are known as cellodextrins; in contrast to long-chain cellulose, cellodextrins are typically soluble in water and organic solvents.
Plant-derived cellulose is usually found in a mixture with hemicellulose, lignin, pectin and other substances, while microbial cellulose is quite pure, has a much higher water content, and consists of long chains.
Cellulose is soluble in cupriethylenediamine (CED), cadmiumethylenediamine (Cadoxen), N-methylmorpholine N-oxide and lithium chloride / dimethylformamide.This is used in the production of regenerated celluloses (as viscose and cellophane) from dissolving pulp.[42]
Chitin
Chitin is one of many naturally occurring polymers. It is one of the most abundant natural materials in the world. Over time it is bio-degradable in the natural environment. Its breakdown may be catalyzed by enzymes called chitinases, secreted by microorganisms such as bacteria and fungi, and produced by some plants. Some of these microorganisms have receptors to simple sugars from the decomposition of chitin. If chitin is detected, they then produce enzymes to digest it by cleaving the glycosidic bonds in order to convert it to simple sugars and ammonia.
Chemically, chitin is closely related to chitosan (a more water-soluble derivative of chitin). It is also closely related to cellulose in that it is a long unbranched chain of glucose derivatives. Both materials contribute structure and strength, protecting the organism.Chitin is a modified polysaccharide that contains nitrogen; it is synthesized from units of N-acetylglucosamine (to be precise, 2-(acetylamino)-2-deoxy-D-glucose). These units form covalent β-1,4 linkages (similar to the linkages between glucose units forming cellulose). Therefore, chitin may be described as cellulose with one hydroxyl group on each monomer substituted with an acetyl amine group. This allows for increased hydrogen bonding between adjacent polymers, giving the chitin-polymer matrix increased strength.
In its unmodified form, chitin is translucent, pliable, resilient, and quite tough. In arthropods, however, it is often modified, becoming embedded in a hardened proteinaceous matrix, which forms much of the exoskeleton. In its pure form, it is leathery, but, when encrusted in calcium carbonate, it becomes much harder. The difference between the unmodified and modified forms can be seen by comparing the body wall of a caterpillar (unmodified) to a beetle (modified).N-Acetylglucosamine (N-acetyl-D-glucosamine, or GlcNAc, or NAG) is a monosaccharide derivative of glucose. It is an amide between glucosamine and acetic acid. It has a molecular formula of C8H15NO6, a molar mass of 221.21 g/mol, and it is significant in several biological systems.[43]
It is part of a biopolymer in the bacterial cell wall, built from alternating units of GlcNAc and N-acetylmuramic acid (MurNAc), cross-linked with oligopeptides at the lactic acid residue of MurNAc. This layered structure is called peptidoglycan.GlcNAc is the monomeric unit of the polymer chitin, which forms the outer coverings of insects and crustaceans.Polymerized with glucuronic acid it forms hyaluronan.
Arabinoxylans
Arabinoxylans are the copolymers of two pentose sugars - arabinose and xylose.
Bacterial polysaccharides represent a diverse range of macromolecules that include peptidoglycan, lipopolysaccharides, capsules and exopolysaccharides; compounds whose functions range from structural cell-wall components (e.g. peptidoglycan), and important virulence factors (e.g. Poly-N-acetylglucosamine in S. aureus), to permitting the bacterium to survive in harsh environments (e.g. Pseudomonas aeruginosa in the human lung). Polysaccharide biosynthesis is a tightly regulated, energy-intensive process and understanding the subtle interplay between the regulation and energy conservation, polymer modification and synthesis, and the external ecological functions is a huge area of research. The potential benefits are enormous and should enable for example the development of novel antibacterial strategies (e.g. new antibiotics and vaccines) and the commercial exploitation to develop novel applications.
Bacterial capsular polysaccharides
Pathogenic bacteria commonly produce a thick, mucous-like, layer of polysaccharide. This "capsule" cloaks antigenic proteins on the bacterial surface that would otherwise provoke an immune response and thereby lead to the destruction of the bacteria. Capsular polysaccharides are water soluble, commonly acidic, and have molecular weights on the order of 100-1000 kDa. They are linear and consist of regularly repeating subunits of one to six monosaccharides. There is enormous structural diversity; nearly two hundred different polysaccharides are produced by E. coli alone. Mixtures of capsular polysaccharides, either conjugated or native are used as vaccines.
Bacteria and many other microbes, including fungi and algae, often secrete polysaccharides as an evolutionary adaptation to help them adhere to surfaces and to prevent them from drying out. Humans have developed some of these polysaccharides into useful products, including xanthan gum, dextran, welan gum, gellan gum, diutan gum and pullulan.
Most of these polysaccharides exhibit interesting and very useful visco-elastic properties when dissolved in water at very low levels. This gives many foods and various liquid consumer products, like lotions, cleaners and paints, for example, a viscous appearance when stationary, but fluidity when the slightest shear is applied, such as when wiped, poured or brushed. This property is referred to as pseudoplasticity, or shear thinning.[44]
Peptidoglycan
Peptidoglycan, also known as murein, is a polymer consisting of sugars and amino acids that forms a mesh-like layer outside the plasma membrane of bacteria (but not Archaea), forming the cell wall. The sugar component consists of alternating residues of β-(1,4) linked N-acetylglucosamine and N-acetylmuramic acid. Attached to the N-acetylmuramic acid is a peptide chain of three to five amino acids. The peptide chain can be cross-linked to the peptide chain of another strand forming the 3D mesh-like layer.
The peptidoglycan layer in the bacterial cell wall is a crystal lattice structure formed from linear chains of two alternating amino sugars, namely N-acetylglucosamine (GlcNAc or NAG) and N-acetylmuramic acid (MurNAc or NAM). The alternating sugars are connected by a β-(1,4)-glycosidic bond. Each MurNAc is attached to a short (4- to 5-residue) amino acid chain, containing D-alanine, D-glutamic acid, and meso-diaminopimelic acid in the case of Escherichia coli (a Gram negative bacteria) or L-alanine, D-glutamine, L-lysine, and D-alanine in the case of Staphylococcus aureus (a Gram positive bacteria). These amino acids, except the L-amino acids, do not occur in proteins and are thought to help protect against attacks by most peptidases
Cross-linking between amino acids in different linear amino sugar chains occurs with the help of the enzyme transpeptidase and results in a 3-dimensional structure that is strong and rigid. The specific amino acid sequence and molecular structure vary with the bacterial species.[45]
LPS
LPS is the major component of the outer membrane of Gram-negative bacteria, contributing greatly to the structural integrity of the bacteria, and protecting the membrane from certain kinds of chemical attack. LPS also increases the negative charge of the cell membrane and helps stabilize the overall membrane structure. It is of crucial importance to gram-negative bacteria, whose death results if it is mutated or removed. LPS is an endotoxin, and induces a strong response from normal animal immune systems. It has also been implicated in non-pathogenic aspects of bacterial ecology, including surface adhesion, bacteriophage sensitivity, and interactions with predators such as amoebae.
LPS acts as the prototypical endotoxin because it binds the CD14/TLR4/MD2 receptor complex, which promotes the secretion of pro-inflammatory cytokines in many cell types, but especially in macrophages. In Immunology, the term "LPS challenge" refers to the process of exposing a subject to an LPS that may act as a toxin.
LPS is also an exogenous pyrogen (external fever-inducing substance).
Being of crucial importance to gram-negative bacteria, these molecules make candidate targets for new antimicrobial agents.[46]
Some researchers doubt reports of generalized toxic effects attributed to all lipopolysaccharides, in particular, for cyanobacteria.It comprises three parts:
N-linked protein glycosylation (N-glycosylation of N-glycans) at Asn residues (Asn-x-Ser/Thr motifs) in glycoproteins.[47]
The term glycan refers to a polysaccharide or oligosaccharide. Glycans usually consist solely of O-glycosidic linkages of monosaccharides. For example, cellulose is a glycan (or, to be more specific, a glucan) composed of beta-1,4-linked D-glucose, and chitin is a glycan composed of beta-1,4-linked N-acetyl-D-glucosamine. Glycans can be homo- or heteropolymers of monosaccharide residues, and can be linear or branched. Glycan may also be used to refer to the carbohydrate portion of a glycoconjugate, such as a glycoprotein, glycolipid, or a proteoglycan.
Glycans can be found attached to proteins as in glycoproteins and proteoglycans. In general, they are found on the exterior surface of cells. O- and N-linked glycans are very common in eukaryotes but may also be found, although less commonly, in prokaryotes.[48]
N-Linked glycans
N-Linked glycans are found attached to the R-group nitrogen (N) of asparagine in the sequon. The sequon is a Asn-X-Ser or Asn-X-Thr sequence, where X is any amino acid except proline and may be composed of N-acetyl galactosamine, galactose, neuraminic acid, N-acetylglucosamine, fructose, mannose, fucose, and other monosaccharides.
Assembly:
In eukaryotes, N-linked glycans are derived from a core 14-sugar unit assembled in the cytoplasm and endoplasmic reticulum. First, two N-acetyl glucosamine residues are attached to dolichol phosphate, a lipid, on the external side of the endoplasmic reticulum membrane. Five mannose residues are then added to this structure. At this point, the partially finished core glycan is flipped across the endoplasmic reticulum membrane, so that it is now located within the reticular lumen. Assembly then continues within the endoplasmic reticulum, with the addition of four more mannose residues. Finally, three glucose residues are added to this structure. Following full assembly, the glycan is transferred en bloc by the glycosyltransferase oligosaccharyltransferase to a nascent peptide chain, within the reticular lumen. This core structure of N-linked glycans, thus, consists of 14 residues (3 glucose, 9 mannose, and 2 N-acetylglucosamine).[49]
Once transferred to the nascent peptide chain, N-linked glycans, in general, undergo extensive processing reactions, whereby the three glucose residues are removed, as well as several mannose residues, depending on the N-linked glycan in question. The removal of the glucose residues is dependent on proper protein folding. These processing reactions occur in the Golgi apparatus. Modification reactions may involve the addition of a phosphate or acetyl group onto the sugars, or the addition of new sugars, such as neuraminic acid. Processing and modification of N-linked glycans within the Golgi does not follow a linear pathway. As a result, many different variations of N-linked glycan structure are possible, depending on enzyme activity in the Golgi.
Functions and importance
N-linked glycans are extremely important in proper protein folding in eukaryotic cells. Chaperone proteins in the endoplasmic reticulum, such as calnexin and calreticulin, bind to the three glucose residues present on the core N-linked glycan. These chaperone proteins then serve to aid in the folding of the protein that the glycan is attached to. Following proper folding, the three glucose residues are removed, and the glycan moves on to further processing reactions. If the protein fails to fold properly, the three glucose residues are reattached, allowing the protein to re-associate with the chaperones. This cycle may repeat several times until a protein reaches its proper conformation. If a protein repeatedly fails to properly fold, it is excreted from the endoplasmic reticulum and degraded by cytoplasmic proteases.
N-linked glycans also contribute to protein folding by steric effects. For example, cysteine residues in the peptide may be temporarily blocked from forming disulfide bonds with other cysteine residues, due to the size of a nearby glycan. Therefore, the presence of a N-linked glycan allows the cell to control which cysteine residues will form disulfide bonds.
N-linked glycans also play an important role in cell-cell interactions. For example, tumour cells make N-linked glycans that are abnormal. These are recognized by the CD337 receptor on Natural Killer cells as a sign that the cell in question is cancerous.
The targeting of degradative lysosomal enzymes is also accomplished by N-linked glycans. The modification of an N-linked glycan with a mannose-6-phosphate residue serves as a signal that the protein to which this glycan is attached should be moved to the lysosome. This recognition and trafficking of lysosomal enzymes by the presence of mannose-6-phosphate is accomplished by two proteins: CI-MPR (cation-independent mannose-6-phosphate receptor) and CD-MPR (cation-dependent mannose-6-phosphate receptor).[50]
O-Linked glycans
In eukaryotes, O-linked glycans are assembled one sugar at a time on a serine or threonine residue of a peptide chain in the Golgi apparatus. Unlike with N-linked glycans, there is as of yet no known consensus sequence. However, the placement of a proline residue at either -1 or +3 relative to the serine or threonine is favourable for O-linked glycosylation.
Assembly
The first monosaccharide attached in the synthesis of O-linked glycans is N-acetyl-galactosamine. After this, several different pathways are possible. A Core 1 structure is generated by the addition of galactose. A Core 2 structure is generated by the addition of N-acetyl-glucosamine to the N-acetyl-galactosamine of the Core 1 structure. Core 3 structures are generated by the addition of a single N-acetyl-glucosamine to the original N-acetyl-galactosamine. Core 4 structures are generated by the addition of a second N-acetly-glucosamine to the Core 3 structure. Other core structures are possible, though less common.
A common structural theme in O-linked glycans is the addition of polylactosamine units to the various core structures. These are formed by the repetitive addition of galactose and
N-acetyl-glucosamine units. Polylactosamine chains on O-linked glycans are often capped by the addition of a sialic acid residue (similar to neuraminic acid). If a fucose residue is also added, to the next to penultimate residue, a Sialyl_lewis_x (SLex)) structure is formed.[51]
Proteoglycans are glycoproteins that are heavily glycosylated. The basic proteoglycan unit consists of a "core protein" with one or more covalently attached glycosaminoglycan (GAG) chain(s). The point of attachment is a Ser residue to which the glycosaminoglycan is joined through a tetrasaccharide bridge (For example: chondroitin sulfate-GlcA-Gal-Gal-Xyl-PROTEIN). The Ser residue is generally in the sequence -Ser-Gly-X-Gly- (where X can be any amino acid residue), although not every protein with this sequence has an attached glycosaminoglycan. The chains are long, linear carbohydrate polymers that are negatively charged under physiological conditions, due to the occurrence of sulfate and uronic acid groups. Proteoglycans occur in the connective tissue.
Proteoglycans can be categorised depending upon the nature of their glycosaminoglycan chains. Proteoglycans can also be categorised by size (kDa). [52]
Certain members are considered members of the "small leucine-rich proteoglycan family" (SLRP). These include decorin, biglycan, fibromodulin and lumican.
↑Ionescu, A.R.; Berces, A.; Zgierski, M.Z.; Whitfield, D.M.; Nukada, T. (2005). "Conformational Pathways of Saturated Six-Membered Rings. A Static and Dynamical Density Functional Study". The Journal of Physical Chemistry A. 109 (36): 8096–8105. doi:10.1021/jp052197t. PMID16834195.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑Biarns, X.; Ardvol, A.; Planas, A.; Rovira, C.; Laio, A.; Parrinello, M. (2007). "The Conformational Free Energy Landscape of b-D-Glucopyranose. Implications for Substrate Preactivation in b-Glucoside Hydrolases". Journal of the American Chemical Society. 129 (35): 10686–10693. doi:10.1021/ja068411o. PMID17696342.{{cite journal}}: CS1 maint: multiple names: authors list (link)
↑ abMorrison, Robert Thornton and Boyd, Robert Neilson. Organic Chemistry (2nd ed.). Allyn and Bacon.{{cite book}}: CS1 maint: multiple names: authors list (link) Library of Congress catalog 66-25695
↑Derek Horton (2008). "The Development of Carbohydrate Chemistry and Biology". Carbohydrate Chemistry, Biology and Medical Applications: 1–28. doi:10.1016/B978-0-08-054816-6.00001-X.













 D-Erythrose
D-Erythrose D-Threose
D-Threose D-Erythrulose
D-Erythrulose





























{kind=link}









